Theory & Best Practices
"See first, think later, then test. But always see first. Otherwise you will only see what you were expecting." - Douglas Adams
The Biological Theory of Relativity
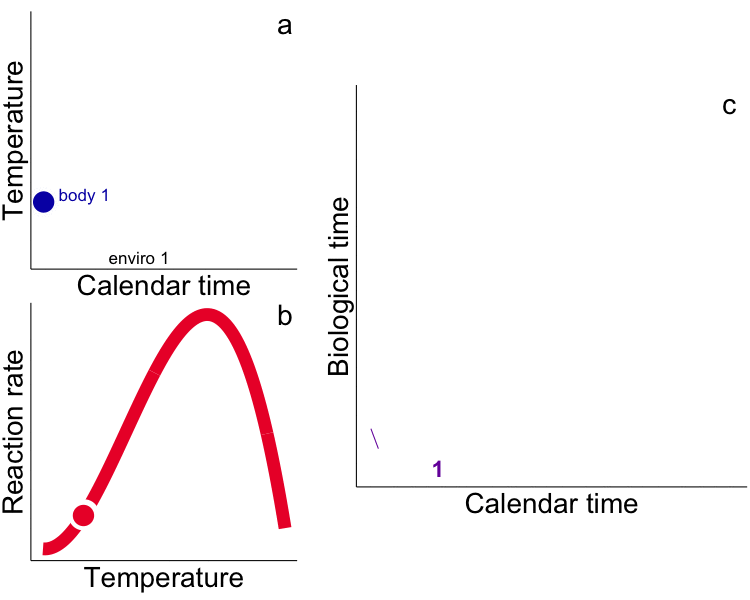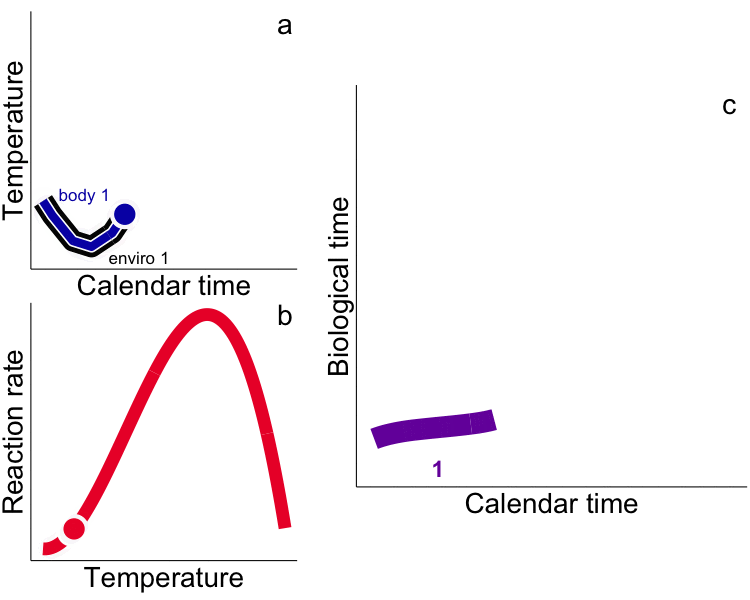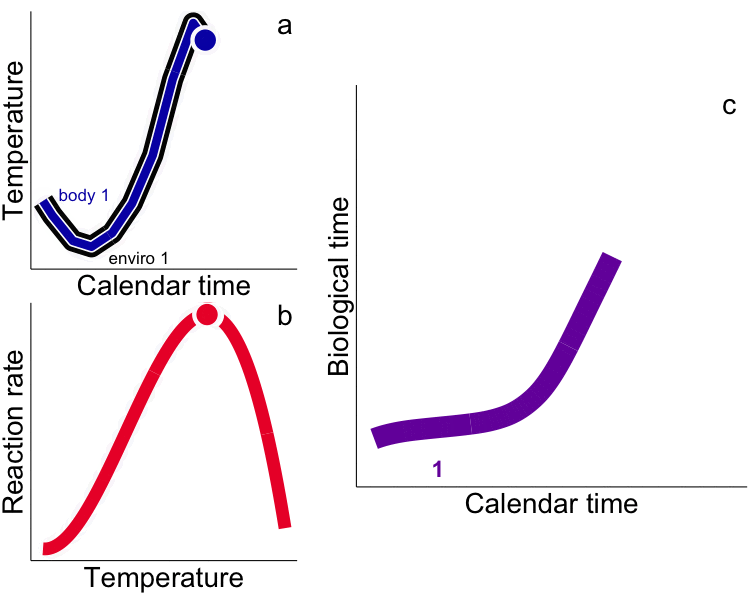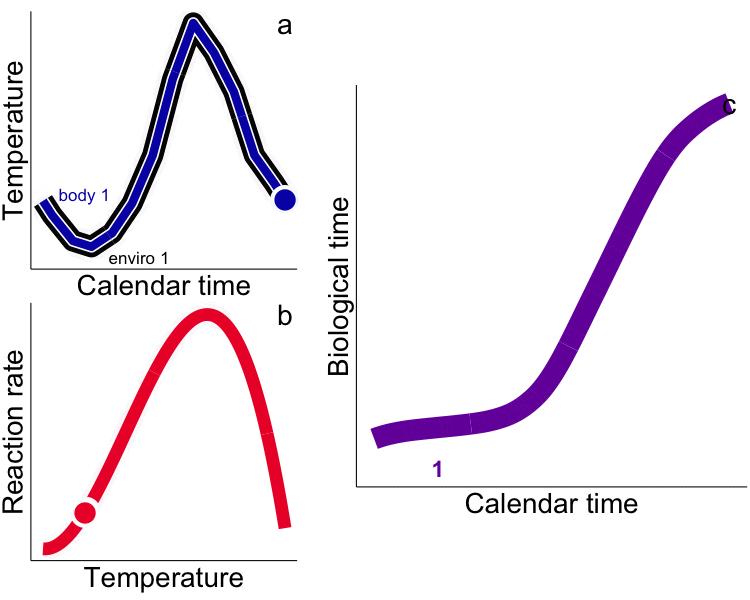
While typical “calendar” time may be appropriate for representing rates for endotherms that experience relatively constant temperatures (e.g. humans), use of calendar time in physiological (e.g. growth) models for ectotherms explicitly ignores time-dependent and physiologically-meaningful temperature variability. Integrated measures of growth (e.g. size) and development (e.g. egg hatch time) in ectotherms depend not only on the time allowed for the process but the temperature. Instead, metrics that incorporate the ectotherm’s thermal history (time spent at temperature) are required, i.e. the time-based integral of the heat available for growth – heat transferred from the environment to the ectotherm – as degree days (DD, or growing degree-days, GDD, ºC∙day). Rates measured in calendar time are aliased by our ‘endothermocentric’ view of time that is physiologically meaningless among ectotherms that have different thermal histories. In the future, temperature by itself, and time by itself should fade away as only a union of the two will preserve an independent reality for ectotherms (cf. Minkowski 1908). We challenge others to test the GDD metric in all aspects of fish and aquatic invertebrate physiology, growth and development. We have used integrated temperature metrics to explain variability and disentangle plastic vs. adaptive variation in a wide range of processes (growth, life history, feeding) and animals (fish and invertebrates) including:
Neuheimer, A.B. The pace of life: Time, temperature and a biological theory of relativity. PREPRINT. bioRxiv. DOI: 10.1101/609446.
Neuheimer, A.B.* & P. Grønkjær. 2012. Climate effects on size-at-age: Growth in warming waters compensates for earlier maturity in an exploited marine fish. Global Change Biology 18:1812-1822. (link)
Neuheimer, A.B.*, C.T. Taggart & K.T. Frank. 2008. Size-at-age in haddock (Melanogrammus aeglefinus) - application of the growing degree-day (GDD) metric. Proceedings of the 24th Lowell Wakefield Fisheries Symposium: Resiliency of Gadid Stocks to Fishing and Climate Change Symposium, 111-123. (pdf)
Neuheimer, A.B.* 2007. Growth in fishes: size-at-age, temperature and food. Ph.D. Thesis, Department of Oceanography, Dalhousie University, Halifax, Canada.
Modelling Strategies & Best Practices
Our lab develops flexible, stochastic, biophysical modelling tools to explain and predict variability in biological systems including recruitment, food consumption, connectivity, etc. Model applications can be found through the links here. In addition, our contributions to modelling strategy and best practices include:
Brander, K., A.B. Neuheimer*, K.H. Andersen & M. Hartvig. 2013. Food for thought: Overconfidence in model projections. ICES Journal of Marine Science 70:1065-1068. (link)
Neuheimer, A.B.*, W.C. Gentleman, P. Pepin & E.J.H. Head. 2010a. How to build and use individual-based models (IBMs) as hypothesis testing tools. Journal of Marine Systems 81: 122-133. (link)
Gentleman, W.C., A.B. Neuheimer* & R.G. Campbell. 2008. Modelling copepod development: Current limitations and a new realistic approach. ICES Journal of Marine Science 65: 399-413. (link)